| DOWNLOAD PDF |
R. L. Gregory, H. E. Ross and N. Moray
Reprinted from Nature, 201, 4925, 1166 (1964) with the kind permission of the Editor
[We now take up a topic concerned with the early evolution of vision.
The nineteenth-century zoologist Seliq Exner reported in 1891 the structure of a copepod living in the bay of Naples, having what turned out to be a most curious eye - possibly a single channel scanning eye, like a simple mechanical television camera, feeding information of spatial structure down a single neural channel in time. Professor J. Z. Young pointed out to me that several compound eyes - especially the eye of Daphnia - are in the kind of 'continual lively motion' reported by Exner for Copilia. So, in short, we decided upon an expedition. This consisted, apart from myself, of Helen Ross and Neville Moray, who though a psychologist (at that time at Sheffield, now in Canada) had a background in zoology.
We had no drawings to go on (though in fact there are early drawings of Copilia) and so we did not know just what to expect. We examined a gallon of water a day, drop by drop, with three microscopes until, though fascinated by what we did see, we began to despair of finding Copilia. Then suddenly there she was! Incredibly beautiful: perfectly transparent - so no veil hid the secrets of her eyes.
This brief investigation led to five years of work, still to be described, on compound eyes of varying complexity and characteristics. This is the work of my students, Stephen Young (now a lecturer in zoology at Imperial College) and Tony Downing who is my colleague. They have developed techniques for studying the vision and behaviour of these creatures. We went on a second expedition in the summer of 1972 - and found Copilia even more fascinating as we came to know her better.]
Exner (1891) described the visual apparatus of the copepod Copilia, which he examined at Naples in the 1880s. Exner's account is discussed by Wilkie (1953), which directed our attention to this eye.
According to Exner, each of the lateral eyes of Copilia has a pair of lenses. The anterior lenses are large, and exceptionally widely separated. The posterior lens of each eye lies a great distance behind the anterior lens - half-way along the extraordinarily transparent body of the animal - so that it is not at first obvious that it has any connection with the eye. The most striking feature reported by Exner is that this second posterior lens was in continual and lively motion, apparently moving across the image plane of the anterior lens. From his description, it appears that there is no retina but rather a single functional receptor unit, transmitting its information to the central brain down a single pathway.
With what we now know about transmitting spatial information by conversion into a time-series by scanning, as in television, it seemed possible that Exner was describing an organism the eye of which works on a principle now very familiar to the engineer. We decided to try to find and examine this animal. This was made possible through the kindness and enthusiasm of Prof. J. Z. Young, and the generous co-operation of the Director and Staff of the Stazione Zoologica di Napoli.
The animals were collected from hauls made at a depth of about 200m in the Bay of Naples. We succeeded in finding 9 living specimens of Copilia, in the course of examining some 14 plankton hauls. Of the nine, eight were female and one male.
Exner's description we found to be accurate with regard to the female specimens, the males being very different. Using high-quality optical microscopes (bright-ground, dark-ground and phase contrast Leitz equipment) we found that the internal structure, muscles, ligaments and the nervous system could easily be observed in the living unstained specimen. In particular, the oscillatory movement of the posterior lens and receptor, as reported by Exner, was readily observable.
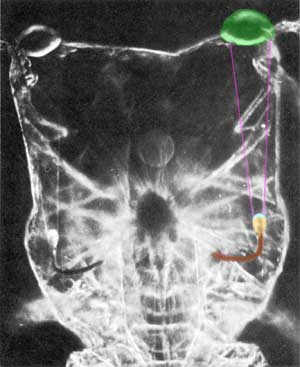
FIG. 1 Photomicrograph of Copilia quadrata showing the whole of the body, from above, but not the tail. The anterior lenses (shaded green) are seen somewhat out of focus; the posterior lenses (shaded pale blue) and the opaque pigment (shaded pale yellow and brown, but actually orange) of the photoreceptors are seen in sharp focus. These 'scan', apparently across the image planes of the anterior lenses. The specimen is living and unstained.
The following remarks apply only to the female.
Copilia quadrata is about 3 mm in total length including the long tail; about 1 mm in width, and about 1 mm in depth through the maximum thickness of the body. The diameter of the anterior lens is about 0.15 mm. The posterior lens is situated about 0.65 mm behind the first, the two being joined by a delicate cone-shaped membrane. The posterior lens, lying deep in the animal, is clearly seen in Fig. 1, which shows the whole of the body but not the tail. The lens is attached to a heavily pigmented bow-shaped, orange coloured structure which contains the photosensitive elements. The optic nerve is clearly seen in the living animal leaving the medial side of this bow-shaped structure, passing thence to the supraosophageal ganglion. It seems that the whole structure is essentially the same as a single ommatidium of a conventional compound eye, except that the distance between the corneal lens and the crystalline cone is vastly increased. The detailed structure of the 'rhabdom' of this 'ommatidium' has been described by Vaissière (1961), whose observations both with optical and electron microscopy show the microstructure to be very similar to the conventional compound eye. There seem to be the usual cluster of receptor cells, generally believed to function as a single unit.
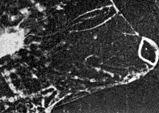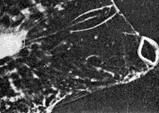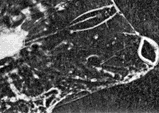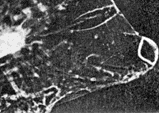 |
FIG. 2 A series of eight consecutive cine-frames of the living animal, during one scan. Photographed at 16 frames/sec. [Reproduced on this web page as an animated GIF file - note that the frame rate is unlikely to be accurate due to limitations in the animation method.]
The anterior ('corneal') lens is rigidly fixed in the strong transparent carapace of the animal. The posterior lens ('the crystalline cone') is suspended in a dynamic system of ligaments and muscles which produce movements of the crystalline cone and its attached photoreceptor across what we assume to be the image plane of the corneal lens, but we were unable to get direct optical evidence for this. The oscillatory movement is 'sawtooth' in form, the receptors moving rapidly towards each other, separating comparatively slowly. The resting state (particularly noticeable in dying animals) is with the receptors farthest apart, when the optical axes of the two eyes are parallel. The axes never converge, and so the 'scanning' (as we are inclined to regard it) can scarcely be associated with range-finder distance vision. The maximum amplitude of the scan is about four times the diameter of the crystalline cone. The scan appeared unrelated to movements of other body structures, which were easy to observe simultaneously. We were able to confirm the independence of the movement of the eye parts by examination of cinematograph film of living specimens. This autonomy seems strong evidence for regarding the movement as scanning. We found the frequency to be very variable (though Exner reports it as constant), but the variability may have been related to the condition of the specimens, which we were unable to keep alive for more than about 12 h. The maximum observed frequency was about 15 scans per sec. in Copilia quadrata, though there appear to be species differences, which will be described in a subsequent report. A single scan is shown in the series of consecutive ciné frames in Fig. 2. We were unable to produce systematic variations in the scan by subjecting the animals to visual stimuli, although there were frequent spontaneous variations in amplitude and in frequency. Even violent changes in illumination (occluding the microscope lamp) produced no related changes in scanning, though the animals would try to avoid too bright a light. We regret that we did not use a red filter while examining them, as the animals are probably insensitive to red light. The maximum intensity they would encounter at the depth we found them is about that of moonlight, but we examined them with many times this intensity.
At this stage we can only speculate as to the place of this eye in the evolutionary sequence. Is it an unsuccessful 'experiment'? Is it a precursor of the compound eye - multiple ommatidia developing to overcome the limited information channel capacity of a neural path? This most curious of eyes seems an ideal target for a microelectrode.
![]()